Těsné křižovatky jsou proteinové sítě. Obepínají endoteliální tkáň střeva, močového měchýře a mozku a kromě stabilizačních funkcí také přijímají bariérové funkce. Poruchy těchto bariérových funkcí mají negativní vliv na různé prostředí těla.
Co je těsná křižovatka?
Každá buněčná membrána obsahuje odlišné proteiny. Jednotlivé membránové proteiny vytvářejí více či méně hustou síť. V této souvislosti je „těsné spojení“ v latině „Zonula occlusionens“ a v angličtině „Tight Junction“ druhem terminálního proužku obsahujícího protein, který například obepíná epiteliální buňky obratlovců a je úzce spojen se sousedními buněčnými vazy.
Úzké spoje těsní mezery mezi buňkami. Odpovídají bariéře difúze. Difúze je látková transportní cesta v těle živých bytostí, která absorbuje jednotlivé molekuly do buněk. Ve formě difúzní bariéry řídí těsné spojení tok molekul do epitelu. Rovněž zabraňují difúzi membránových složek z apikální do laterální oblasti a naopak. Prostřednictvím druhé funkce udržují polaritu epiteliálních buněk.
Úzké křižovatky obepínají ledviny, močový měchýř a střevní epitel. Kromě toho jsou funkční složkou tzv. Hematoencefalické bariéry a zajišťují, aby látky z krve nemohly difundovat do mozkové tkáně. Finální hřebeny vyrobené z membránových proteinů mohou obsahovat různé proteiny. Pravděpodobně ještě ne všechny jsou známy.
Anatomie a struktura
Nejdůležitějšími membránovými proteiny v těsných spojeních jsou claudiny a okluzin. Claudiny byly dokumentovány ve více než 20 různých obratlovcích. Všechny integrální membránové proteiny mají síťové uspořádání a spojují membrány několika buněk v kontaktu hlava-hlava. Anatomii tvoří vodní póry.
Složení obsažených membránových proteinů se liší od epitelu k epitelu a závisí na funkčních požadavcích těsných spojení. Například claudin 16 v renálním epitelu se podílí na příjmu iontů Mg2 + ledvinami do krve. Těsné křižovatky vytvářejí sítě různé těsnosti v závislosti na úkolu a epitelu. Membránové proteiny volně sedí ve střevě. Bariéra krev-mozek tvoří relativně těsnou bariéru.
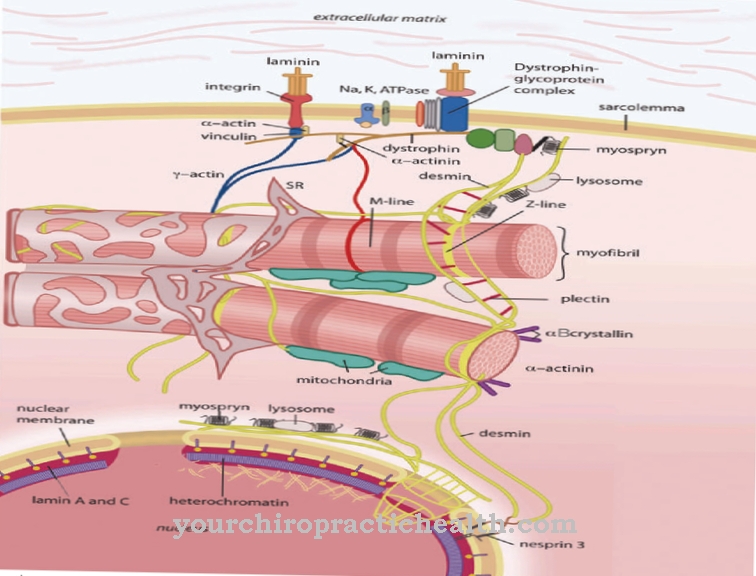
Těsnost sítě koreluje s propustností. Proteinová síť se skládá z úzkých vláken. Zejména se extracelulární oblasti jednotlivých proteinů spojí a vytvoří buněčné spojení. Vnitrobuněčné oblasti závisí na cytoskeletu buněk. Těsné křižovatky obklopují obvod buňky epitelu jako pás a tak se hnízdí proti struktuře epiteliálních buněk.
Funkce a úkoly
Těsné křižovatky jsou primárně difúzní bariérou. Tato funkce může zadržovat molekuly zcela z intracelulárního prostoru, nebo může být spojena se selektivní permeabilitou (semipermeabilitou) pro molekuly určité velikosti. Síť těsných křižovatek je díky své funkci difúzní bariéry předpokladem transcytózy. Paracelulární difúze molekul nebo iontů epitelovým prostorem je zabráněno těsnými spojeními. Koncové pásky zároveň zabraňují vytékání tělních tekutin.
Membránové proteiny těsných spojení také chrání organismus před napadením mikroorganismy, a tak také vytvářejí bariéru pro živé vetřelce. Kromě funkce bariéry mají těsné křižovatky také tzv. Plotovou funkci. Proteinová síť zabraňuje pohybu jednotlivých složek membrány a udržuje tak buněčnou polaritu epitelu. Epitel je sítěmi rozdělen na apikální a bazální oblasti. Apikální buněčná membrána epitelu má jinou biochemii než bazolaterální buněčná membrána. Úzké křižovatky pomáhají udržovat tyto rozdíly v biochemickém prostředí a umožňují tak řízený transport látek.
K těmto funkcím jsou přidány mechanické funkce. Například těsné spojení také slouží ke stabilizaci sestav epitelových buněk. Spojují buňky cytoskeletu navzájem a zajišťují tkáňovou statiku epitelu. Propustnost mezi epitelovými buňkami podléhá dočasným změnám. Epitel je tedy schopen reagovat na zvýšené požadavky na paracelulární transport. K tomuto účelu se klaudiny a okluziny "těsných spojení" spojují s proteiny intracelulární membrány, které se připojují k aktinovému cytoskeletu.
Zde najdete své léky
➔ Léky na svalovou slabostNemoci
Úzké spoje mohou podléhat změnám ve struktuře kvůli mutacím, a tak ztratit své funkce. Claudin 16 proteinových sítí v epitelu ledvin není přítomen v požadované formě po mutacích v genu kódujícím protein. Takové mutace mohou vést ke ztrátě Mg2 +.
Kvůli ztrátě bariérové funkce je příliš málo Mg2 + iontů absorbováno z ledvin do krve a příliš mnoho je vylučováno močí. Nemoci mohou také ovlivnit „zonula occlusionens“. To platí zejména o mozku. Krev-mozková bariéra je přirozená difúzní bariéra mezi krví a mozkem, která udržuje prostředí mozku. Poruchy hematoencefalické bariéry se vyskytují například v souvislosti s roztroušenou sklerózou. Nemoci, jako je diabetes mellitus, však mohou narušit hematoencefalickou bariéru. Ochranný účinek bariéry je také ztracen při různých zraněních mozku a degenerativních onemocněních.
U roztroušené sklerózy je to opakující se zánět mozku, který má škodlivý vliv na těsné křižovatky. Buňky vlastní imunitní obrany těla překonávají hematoencefalickou bariéru jako součást autoimunitního onemocnění.Při ischemické cévní mozkové příhodě se dokonce rozpadají složky těsných spojení v hematoencefalické bariéře. Tento typ mrtvice je doprovázen prázdnotou v mozku, která je poté naplněna krví. Endothelia hematoencefalické bariéry se mění ve dvou fázích.
Protože patologický proces uvolňuje oxidanty, proteolytické enzymy a cytokiny, mění se propustnost hematoencefalické bariéry. Edém se vyvíjí v mozku. Aktivované leukocyty pak uvolňují tzv. Matricové metaloproteázy, které v těsných spojeních rozkládají bazální laminu a proteinové komplexy.


.jpg)




.jpg)