Ukončení je konečná fáze replikace DNA. Předchází mu iniciace a protažení. Včasné ukončení replikace může vést k expresi zkrácených proteinů a tím k mutaci.
Co je ukončení?

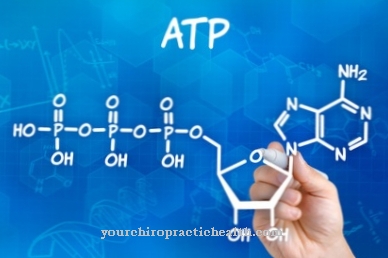
Během replikace nebo reduplikace je DNA genetického informačního nosiče rozmnožena v jednotlivých buňkách. Duplikace probíhá podle polokonzervativního principu a obvykle vede k přesné duplikaci genetické informace. Replikace je spouštěna během fáze syntézy, před fází mitózy, a tak probíhá před dělením buněčného jádra.
Na začátku replikace je dvouřetězcový řetězec DNA rozdělen do jediných řetězců, na nichž jsou vytvořeny nové komplementární řetězce. Každý řetězec DNA je určen základní sekvencí opačného řetězce. Replikace DNA probíhá v několika fázích. Ukončení je třetí a tedy poslední fáze replikace. Ukončení předchází zahájení a prodloužení.
Synonymním termínem pro vyjádření ukončení v této souvislosti je označení Ukončovací fáze. Ukončení zde znamená „přerušení“ nebo „ukončení“. Během ukončení se nově vytvořené vlákno mRNA oddělí od skutečné DNA. Práce DNA polymerázy pomalu končí. Ukončení replikace DNA by nemělo být zaměňováno s ukončením replikace RNA.
Funkce a úkol
Ve fázi iniciace replikace dochází především k regulaci replikace. Je stanoven počáteční bod replikace a dochází k tzv. Aktivaci. Po iniciaci začíná polymerizace, během níž probíhá prodlužovací fáze. Enzym DNA polymeráza rozděluje komplementární řetězce DNA na jednotlivé řetězce a čte základny jednotlivých řetězců jeden po druhém. V této fázi dochází k polospojitému zdvojnásobení, které zahrnuje další fázi aktivace.
Teprve po zahájení a prodloužení následuje fáze ukončení v rámci replikace. Ukončení se liší od formy života k formě života. V eukaryotech, jako jsou lidé, je DNA strukturována do kruhu. Zahrnuje také terminační sekvence, které odpovídají dvěma různým sekvencím, z nichž každá je relevantní pro replikační vidlici.
Ukončení obvykle není vyvoláno speciálními mechanismy. Jakmile dvě replikační vidlice běží společně nebo končí DNA, replikace se v tomto bodě automaticky ukončí. Replikace je ukončena automatickým mechanismem.
Terminační sekvence jsou kontrolní prvky. Zajišťují, aby replikační fáze dorazila ke konkrétnímu koncovému bodu kontrolovaným způsobem, a to i přes různé rychlosti replikace ve dvou replikačních vidlicích. Všechna terminační místa odpovídají vazebným místům pro protein Tus, „látka využívající terminus“. Tento protein blokuje replikativní helikázu DnaB a tak zastavuje replikaci.
V eukaryotech zůstávají replikované prstencové řetězce navzájem propojeny i po replikaci. Připojení odpovídá koncovému bodu. Teprve po dělení buněk jsou odděleny různými procesy a lze je tedy rozdělit. Zdá se, že spojení, které přetrvává až po dělení buněk, hraje roli v řízené distribuci.
Při konečné separaci prstenů DNA hrají roli dva hlavní mechanismy. Enzymy, jako je topoisomeráza typu I a typu II, jsou zapojeny do separace. Nakonec pomocný protein rozpoznává stop kodon během terminace. To způsobí, že polypeptid odpadne z ribozomu, protože není k dispozici žádná t-RNA s vhodným antikodonem pro stop kodon. Ribozom se nakonec rozpadne na dvě podjednotky.
Nemoci a nemoci
Všechny procesy pro zdvojování genetického materiálu ve smyslu replikace jsou komplikované a vyžadují vysoké výdaje látek a energie v buňce. Z tohoto důvodu mohou snadno nastat spontánní chyby replikace. Pokud se genom změní spontánně nebo je navenek zvenčí, mluvíme o mutacích.
Chyby replikace mohou mít za následek chybějící základny, mohou být spojeny se změněnými základnami nebo mohou být výsledkem nesprávného párování bází. Kromě toho může delece a inzerce jednoho nebo více nukleotidů do dvou řetězců DNA vést k chybám replikace. Totéž platí pro pyrimidinové dimery, konce vláken a chyby zesíťování v řetězcích DNA.
V případě chyby replikace jsou k dispozici samostatné opravné mechanismy. Mnoho z uvedených chyb je v maximální možné míře korigováno DNA polymerázou. Přesnost replikace je relativně vysoká. Míra chybovosti je pouze jedna chyba na nukleotid, což je způsobeno různými kontrolními systémy.
Například, kontrolní mechanismus eukaryotických buněk je známý jako nesmyslem zprostředkovaný rozpad mRNA, který dokáže rozpoznat nežádoucí stop kodony v mRNA, a tak zabránit zkráceným proteinům v nalezení exprese.
Předčasné stop kodony v mRNA jsou způsobeny genovými mutacemi. Takzvané nesmyslové mutace nebo alternativní a vadné sestřihování mohou vést ke zkrácení proteinů, které jsou ovlivněny funkčními ztrátami. Kontrolní mechanismy nemohou vždy opravit chyby.
Autozomálně recesivní zděděná porucha β-thalassémie přichází ve třech různých formách: první je homozygotní thalassémie, vážné onemocnění, které je způsobeno vaší nesmyslnou mutací. Heterozygotní thalassémie je mírnější onemocnění, u kterého jsou nesmyslné mutace nalezeny pouze v jedné kopii genu β-globinu. Mechanismem nesmyslem zprostředkovaného rozpadu mRNA může být mRNA vadného genu degradována do té míry, že jsou exprimovány pouze zdravé geny.
Při heterozygotní thalassemii a tedy střední formě onemocnění je nesmyslná mutace v posledním mRNA exonu, takže kontrolní mechanismy nejsou aktivovány. Z tohoto důvodu se kromě zdravého β-globinu tvoří také zkrácený β-globbin.Erytrocyty s vadným β-globinem hynou.
Dalším příkladem selhání kontrolního mechanismu je Duchenneova svalová dystrofie, která je také způsobena nesmyslnou mutací v mRNA. V tomto případě kontrolní mechanismus rozkládá mRNA, ale tak způsobuje úplnou ztrátu takzvaného dystrofinového proteinu.




.jpg)



.jpg)





.jpg)
.jpg)

.jpg)